 |
| Yacimientos neandertales. O. Jöris et al (2011). |
Múltiples yacimientos en Europa y Asia cercana pero
no en África.
Los
fósiles de la última parte del Pleistoceno medio (desde hace unos 200.000 años)
ya pueden ser considerados, a todos los efectos, verdaderos y completos neandertales.
A
partir de hace 127 ka, los fósiles neandertales empiezan a ser abundantes en los
yacimientos europeos, y se encuentran también en Asia central y Oriente Próximo,
lugares a donde emigraron desde Europa. Fósiles tan emblemáticos como Le Moustier,
que da nombre al Musteriense, Guattari 1 (Monte Circeo) o La Chapelle-aux-Saints,
vivieron en Europa hace menos de 60.000 años. Estos neandertales tienen algunas
características nuevas con respecto a los anteriores, y son llamados a menudo "neandertales
clásicos". Los últimos neandertales vivieron en Europa hasta hace unos 30.000
años, e incluso algo menos, antes de extinguirse.
 |
| Análisis de componenetes de la morfología de la superficie articular de la escápula del esqueleto de Altamura. Martina Lari et al (2015) |
Anteriores a hace 134 ka.
Apidimia 2 (Península Mani, Sur de Grecia; Harvati et al, 2019) preserva el esqueleto facial y una gran parte de la caja craneana, pero sin el hueso occipital. Bartsiokas et al (2017) han datado un fragmento de hueso craneal mediante ablación láser en series de uranio, obteniendo una edad mínima de hace 160 ka. lo que hace de Apidimia 2 el fósil más temprano con una clara morfología neandertal.
Martina Lari et al (2015) han datado y realizado una caracterización preliminar, además de un análisis del ADNmt, del esqueleto fósil hallado en 1993 en el karst de la Cueva de Lamalunga, cerca de Altamura, en el sur de Italia y que en su mayor parte continúa atrapado en las formaciones de calcita de la cueva. De acuerdo con los resultados, se trata de un neandertal datado entre 172 ± 15 ka a 130,1 ± 1,9 ka.
En la Cueva Bajondillo (Málaga) hay evidencias de neandertales y del consumo de mariscos, con una datación de 150 ka (Miguel Cortés-Sánchez y otros, 2011).
En la Cueva Bajondillo (Málaga) hay evidencias de neandertales y del consumo de mariscos, con una datación de 150 ka (Miguel Cortés-Sánchez y otros, 2011).
En Tossal de la Font (Castellón) se encontraron un fragmento
de húmero y otro de pelvis.
Francia
- Le Lazaret. Paietal de inmaduro, con cicatriz de trauma y meningioma postraumático; un incisivo lacteal y un canino adulto. Asociado a industria modo 2 evolucionado. 0,16 mda.
- Grotte Suard. La Chaise. Pates de un cráneo adulto, otro infantil y su mandíbula, tres dientes. Asociados a industria tipo 2 evolucionado, con lascas Levallois. 0,13 mda.
- Orgnac 3 (Combier, 1967). Siete dientes asociados a unos pocos bifaces.
- Payre G y F (Moncel et al, 2002). Parietal y ocho dientes asociados a unos pocos bifaces.
- Abri Moula c. XVI. Un infante.
Iberia, Pirineos y Mediterráneo.
|
Europa Occidental
|
Europa Central, Norte y Oriental y Oriente Próximo
|
Asia
|
|
170-74 ka
|
Altamura
Crombe Grenal
Guattari
Apidimia
Kaprina
|
Tabun
|
||
74-59 ka
|
La Quina
|
Subalyuk
Kebara |
Shanidar
|
|
59-45 ka
|
El Sidrón
La Ferrassie Sima de las Palomas del Cabezo Gordo |
Khulna
|
||
45-35,8 ka
|
Le Moustier
La Chapelle-aux-Saints
|
Feldhofer
Vindija |
||
35,8-27 ka
|
Saint-Césaire
Gorham
|
Spy d’Orneau
|
 |
| Localización y estratigrafía del yacimiento de Payre. Michaela Ecker et al, 2013. |
Datados en el gran interglacial del periodo OIS5 (134.000-74.000 años).
Son raros y dispersos.
Moula-Guercy. Fragmentos craneales. Restos poscraneales.
Artenac (Charente). Fragmentos craneales.
Abri Moula c. XV (Defleur et al 1999). 90 restos humanos.
Payre E y D. Dos dientes.
Abri des Pêcheurs. Siete dientes.
Abri du Maras, nivel 5.
La Chaise. El sitio se compone de tres cuevas: Bourgeois-Delauny, Suard y Duport. Descubierta en la década de 1850. Explorada sistemáticamente en 1930. Las cuevas fueron excavadas luego intermitentemente hasta finales de 1970.
Próximo Oriente
Tabün. Pelvis de hembra neandertal más completo del registro fósil.
Francia
Cueva de Crombe Grenal, Dordoña, Francia. Ocupada por neandertales entre 115-50 ka. Excavada en 1950-1960 por François Bordes.- Según el estudio de Phillip Chase de los huesos de animales hallados en el yacimiento, los neandertales cazaban ciervos y renos, de uno en uno, y aprovechaban la carroña de caballos y bóvidos.
- Indicios de utilización simbólica del esqueleto de rapaces diurnas (Morin y Laroulandie, 2012).
Moula-Guercy. Fragmentos craneales. Restos poscraneales.
Artenac (Charente). Fragmentos craneales.
Abri Moula c. XV (Defleur et al 1999). 90 restos humanos.
Payre E y D. Dos dientes.
Abri des Pêcheurs. Siete dientes.
Abri du Maras, nivel 5.
La Chaise. El sitio se compone de tres cuevas: Bourgeois-Delauny, Suard y Duport. Descubierta en la década de 1850. Explorada sistemáticamente en 1930. Las cuevas fueron excavadas luego intermitentemente hasta finales de 1970.
- De particular importancia es el. Abri Bourgeois-Delauny que contenía los restos de 23 fósiles de adultos e infantil. El material fósil se compone principalmente de un cráneo parcial, fragmentos de mandíbula y dientes.
- El material del Abri Suard consiste en 52 adultos fragmentarios y bebés. Sobresalen una gran parte de una bóveda craneal, un hueso occipital, fragmentos de mandíbula y dientes asociados.
Alemania
Taubach.Salzgitter.
Ehringsdorf. Cráneo.
 |
| Guattari |
Italia
- Grotta Guattari (Monte Circeo). Un cráneo y dos mandíbulas. El sitio parece haber sido el resultado de la acción de las hienas.
Eslovaquia.
Gánovce. Fragmentos de cráneo, molde endocraneano y otros de poscraneanos.Grecia.
Acantilado de Apidima.- Apidima I. Medio cráneo.
- Apidima II. Varios fósiles humanos con partes del cráneo facial. Primitivos.
Croacia.
El registro de Kaprina, datado en 130 ka, contiene cientos de fósiles de unos ochenta individuos. Primitivos. Se infieren prácticas caníbales. Es un refugio de roca arenisca descubierto por Dragutin Gorjanovic-Kramberger en 1899 cerca de Krapina (Croacia). Excavado entre 1899-1905, este yacimiento proporcionó también más de 800 herramientas líticas y restos de fauna. Salvo las garras y una falange de pigargo, la fauna no muestra signos de manipulación humana, incluyendo otros restos de ave. Los hallazgos se custodian en el Museo de Historia Natural de Croacia (Zagreb).Cuando Gorjanovic publicó sus hallazgos en 1906, descartó que los neandertales fueran una patología de los humanos modernos. El gran número de individuos representados ha permitido estimar el dimorfismo sexual y el rango de variación dentro de la población. Kaprina 5 (Caspari y Radovcic, 2006) muestra una gran robustez en contraste con otros cráneos de este yacimiento.
- Janet Monge et al (2013) describen una displasia fibrosa en K120,71 (fragmento de costilla izquierda).
 |
| Kaprina C |
Península Ibérica.
Lezetxiki, Mondragón, Guipúzcoa. Primer molar y segundo premolar.Bolomor, Tavernes, Valencia. 5 dientes y fragmento de fémur.
Tossal de la Font, Castellón. Dos restos poscraneales.
Cova Negra de Bellús, Xátiva, Valencia. Nivel 33. Fragmento mandibular.
Cova Foradá, Oliva, Valencia. Nivel 48. Hogar con restos de lepóridos y otra fauna allí consumida.
Pinilla del Valle, Madrid. La Niña de Lozoya. Cuatro dientes de un neandertal de corta edad, con indicios de enterramiento intencionado (2011). 20 pares de astas de uros cerca de fogatas. En el mismo valle, Abrigo de Navalmaíllo y Cueva de la Buena Pinta.
Cueva Corazón, Cañón de la Horadada (Mave, Palencia). Yacimiento musteriense. los restos indican diversas actividades ligadas al desollado, el desarticulado y el descarnado. Junto al aprovechamiento cárnico, la médula de los animales también sería explotada como muestran las marcas de percusión y los patrones de fracturación.
Los Casares, Riba de Saelices, Guadalajara. Metacarpiano. Extraviado.
Cueva de Valdegoba, Huérmeces del Cerro, Burgos. Mandíbula, varios dientes, falange de la mano, dos metatarsianos y otros, pertenecientes al menos a cinco individuos distintos.
Carihuela, Granada. Carihuela-3. Hueso frontal de un niño, depositado por algún depredador.
Cova Eirós, Triacastela, Lugo. Artefactos que fueron utilizados como armas de caza. Otros útiles se emplearon para despiece. Otra parte del utillaje sirvió para cortar y tallar madera, posiblemente para fabricar los mangos de las armas de caza. Por último, otros artefactos se usaron para preparar pieles secas que supuestamente sirvieron para confeccionar vestimentas.
Figueira Brava, Portugal. Aprovechamiento de recursos costeros en un 50% de la dieta, grandes mamíferos, plantas del clima mediterráneo y pequeñas presas. Zilhao et al, 2020.
Datados en la glaciación del periodo OIS4 (74.000-59.000).
Francia.
Regourdou. Sepultura datada hace 70.000 años. Asociada a hallazgos líticos musterienses de tipo Ferrassie.- Regourdou 1. Esqueleto parcial muy completo.
- H5. Esqueleto incompleto de adulto.
- H9. Mandíbula de adulto.
- H18. Cráneo infantil.
Abri des Pechêurs. Seis dientes.
Hungría.
Subalyuk. Esqueleto poscraneal, mandíbula de adulto y cráneo infantil, con industria musteriense.Chequia.
Ochoz. Mandíbula de adulto, molar y fragmentos de huesos poscraneales.Italia.
Caverne delle Fate, Savona, Liguria. Frontal, occipital, zigomático, fragmentos de mandíbulas y dientes sueltos.Próximo Oriente.
"Shanidar Site". Ralph Solecki descubrió 9 esqueletos de neandertal entre 1953 y 1960 en la cueva de Shanidar (Irak). Su edad está comprendida entre los 70 y 40.000 años. Restos de otro individuo fueron recuperados posteriormente. En 2018-2019 se informó del descubrimiento de los restos de otros individuos.- Uno de ellos, Shanidar 4, fué enterrado con una ofrenda de flores (aunque algunos lo discuten) En 1971 Solecki escribió el libro "Shanidar, the First Flower People", en el que trataba de quitar los estereotipos de semi-humanos brutos acuñado para los neandertales. Los restos hallados en 2018 corresponden a dos individuos superpuestos con una pidera encima colocada posiblemente de forma intencionada.
- Otro esqueleto, Shanidar 1, estaba sordo, parcialmente ciego, mutilado de un brazo y su marcha era anormal (Trinkaus y Villotte, 2017); si sobrevivió hasta los cuarenta años de edad, aproximandamente, fue porque le ayudaron, lo cual demuestra una compleja organización social. En la muestra, hay una pequeña concavidad correspondiente a la fosa canina y el malar está ligeramente angulado bajo el ángulo inferoexterno de la órbita.
- Shanidar 5 es un esqueleto parcial con evidencia de traumatismo craneal; en 2015-2016 se recuperaron fragmentos que podrían pertenecer al mismo individuo (Pomeroy et al, 2017).
En relación a los hallazgos europeos, los de Asia Occidental muestran una frente más vertical, neurocráneo más elevado y occipitales menos prominentes; debido a la altura de la bóveda, el contorno transversal es diferente; la región central de la cara, aunque bien desarrollada, no presenta la misma oblicuidad hacia atrás y hacia fuera. El arco cigomático es más grueso y su raíz se encuentra un poco más arriba en relación con el meato auditivo. Las apófisis mastoideas son más prominentes (Vandermeersch y M. D. Garralda, 2011).
Estas autapomorfias nos invitan a investigar cuando se produjo la divergencia entre las poblaciones europeas y asiáticas.
Península Ibérica.
Covalejos, Cantabria. Dos dientes de muda.Cova Negra de Bellús, Xátiva, Valencia. Niveles 20-17. Restos de 7 individuos (4 niños, 1 adolescente y 1 adulto). Cráneo 4, radio y fragmento de peroné, con un hogar a techo.
 |
| Planta y zonificación de El Sidrón |
Datados en el tramo templado OIS 3c (59.000-45.000 años).
La Ferrassie (Francia) es muy conocido por el hallazgo de los restos de dos esqueletos adultos de diferente sexo y unos cinco infantiles (tres niños, un recién nacido y un feto), que se empezaron a desenterrar por Capitan y Peyrony a principios de 1900. Los restos infantiles fueron publicados por Jean-Louis Heim al principio de los años 1980. Debido a su composición, el hallazgo permitió las primeras inferencias sobre el dimorfismo sexual y la ontogenia de la especie. Hay evidencias de enterramiento intencional. Se han datado en MIS 3, entre hace 54 ± 3 y 40 ± 2 ka (Guillaume Guérin et al, 2015).- La Ferrassie 1. 1909. Esqueleto de adulto, incluyendo el cráneo neandertal más completo hallado hasta el momento. Moño occipital, frente inclinada, grandes arcos superciliares y cara muy grande. Aunque estas características son variables en otros neandertales clásicos, en La Ferrassie 1, se muestran todas en un grado significativo. Rompió la clavícula y el fémur, que curaron con secuelas; padecía escoliosis; lesiones en las costillas probablemente relacionadas con una infección pulmonar que pudo causar su muerte; por todo ello, requirió de cuidados (Gómez Olivencia et al, 2018).
- La Ferrassie 2. 1910. Cráneo y esqueleto incompleto de hembra.
- La Ferrassie 3. 1912. Esqueleto infantil casi completo.
- La Ferrassie 4 y 4bis. 1912. Esqueleto infantil.
- La Ferrassie 5. 1920. Feto de siete meses.
- La Ferrassie 6. 1921. Infante de 3-5 años. El cráneo estaba separado más de 1 m.
- La Ferrassie 8. 1970. Esqueleto parcial de un infantil de unos dos años de edad, descubierto en 1970, aunque la mayor parte de los restos fueron recuperados en 1973 por el equipo de H. Delporte. En 2015 se ha publicado el redescubrimiento de restos adicionales en el Museo Nacional de Historia Natural de Francia y en el Musée d'Archéologie Nationale et Domaine National de Saint-Germain-en-Laye (Asier Gómez-Olivencia, Isabelle Crevecoeur y Antoine Balzeau, 2015).
La Cotte de St. Helier en la isla de Jersey
Khulna (Chequia). Cráneos, mandíbula y poscraneales de al menos 11 individuos. Maxilar derecho, pragmento de parietal derecho y dientes aislados datados en 45,6 ka.
Carihuela-2. Nivel VI.
Sima de las Palomas, Cabezo Gordo, Torre Pacheco, Murcia. 89 restos. Tres esqueletos neandertales superpuestos: dos adultos (SP92 y SP96) y un infantil (SP97) asociados a herramientas musterienses y fauna (algunos huesos, carbonizados). SP92 podría estar relacionado con SP1. SP96 Paloma se recuperó en 2006-2007 y constituye el esqueleto neandertal más completo de todo el litoral mediterráneo europeo, con la pelvis femenina neandertal más completa conocida. Hay indicios de deposición intencional.
Cueva del Gegant de Sitges, Barcelona. Mandíbula.
Cova de les Teixoneres, Moiá, Barcelona. Dientes de infantil.
La Güelga, sector oriental del Macizo Asturiano.
Boquete de Zafarraya, Alcaucín, Málaga. 3 mandíbulas, escápula, húmero, pubis, 3 fémures, tibia y diente lacteal de nueve individuos.
Vanguard Cave, Gibraltar. Canino superior derecho deciduo.
Jarama VI.
Valdegoba. 48,5 ka.
El Sidrón, Piloña, Asturias. En contexto secundario en un sistema kárstico (M. de la Rasilla et al, 2014; Dataciones: R. E. Wood et al, 2013; M. de la Rasilla et al, 2013). Más de 2.400 fósiles (más de 200 dientes) de al menos 13 individuos: uno infantil (2-3 años) otro infantil casi completo (6-7 años, hijo de una de las mujeres del grupo y hermano del anterior) con molares de leche e incisivos definitivos incipientes), otro juvenil posiblemente masculino, dos de adolescentes varones, siete de jóvenes adultos (tres hembras, tres machos y uno indefinido). Los infantiles, sin dientes. Asociados a 415 artefactos líticos y 51 restos faunísticos. En 2013 se hallaron un fragmento craneal y un maxilar de adulto al que faltan los molares de uno de los lados.
- Se infiere que yacían enterrados próximos a una entrada de la cueva y fueron arrastrados con el sedimento que les envolvía. Pueden ser considerados miembros de una misma población o grupo familiar.
- El análisis genético de la muestra indicó la presencia de una pequeña comunidad patrilocal con baja diversidad genética (Sergi Castellano et al, 2014). Tres varones pertenecen al mismo linaje y tres hembras a linajes diferentes. Con evidencia genética y esquelética de endogamia, podrían ser representativos del inicio del colapso demográfico de este fenotipo (Ríos et al, 2019).
- Dos atlas presentan un defecto congénito muy raro en los humanos modernos en el arco anterior y posterior del atlas, una condición en la mayoría de los casos no se asocia con ninguna manifestación clínica (Ríos et al, 2015). Esta observación, junto con la evidencia informada anteriormente de retención del canino deciduo mandibular en dos de diez denticiones (Dean et al, 2013), apoya la evidencia genética de que estos neandertales constituían un grupo con estrechas relaciones genéticas.
- Cráneo plano y bajo. Capacidad encefálica media 1.500 cc. Frente baja e inclinada hacia atrás. Arcos superciliares grandes. Abertura nasal muy ancha. Pómulos hacia atrás y proyección de la cara media. Carencia de fosa canina. Moño occipital.
- Dientes de tamaño grande. Los incisivos en pala, muy curvados y con fuertes tubérculos en los bordes laterales internos. Los premolares, asimétricos, con cúspides accesorias y crestas transversas. Algunos molares taurodontos.
- Tres mandíbulas están completas o casi. El cuerpo mandibular es alto y grueso. Trígono mentoniano, sin escotadura. Espacio retromolar corto comparado con otros neandertales.
- La comparación con otros neandertales sugiere variabilidad racial y geográfica.
- Cálculos en los dientes de todos los adolescentes y adultos; absceso apical en la mandíbula 2, indicador de periodontitis; en la mitad de las piezas dentarias se observan surcos en sentido vertical en la cara anterior de un diente y la posterior del contiguo, de etiología problemática; hipoplasia del esmalte dentario que señala deficiencia nutricional; sarro; descascarillado. Hipoplasias relacionadas con carencias alimenticias. Evidencias de uso de palillos y de utilización de los dientes como una tercera mano. Dos individuos relacionados vía materna, presentan indicios de caninos deciduos retenidos en la mandíbula izquierda (MC Dean et al, 2013).
- Los huesos presentan marcas y cortes de origen antrópico, pero no en todos los casos, las de los huesos largos como para desarticularlos rompiendo ligamentos. Fractuas indicativas de extracción de médula y sesos. Parece fuera de duda la práctica de canibalismo alimentario.
- En una adulta joven, se han hallado restos de hierbas medicinales amargas: manzanilla y milenrama, lo que sugiere automedicación gracias al conocimiento de las cualidades medicinales.
- Laura T. Buck y Chris B. Stringer (2013) exponen una hipótesis alternativa, basada en la etnografía moderna: los rastros de plantas medicinales en los dientes de los neandertales se deben al consumo de estómagos de hervíboros.
- Conjunto lítico musteriense asociado, pero poco numeroso.
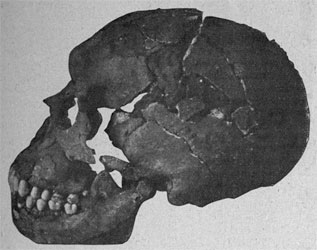 |
| Le Moustier 1. Fotografía de Otto Hauser. |
Datados en el intervalo frío OIS 3b
(45.000-35.800 años)
Francia.
- Le Moustier.
- Le Moustier 1. Restos perdidos en un bombardeo de Berlín durante la segunda guerra mundial con expceción de un esqueleto de infantil masculino datado en 42,5 ka.
- Le Moustier 2. Bebé de unos 9 meses exhumado por Denis Peyrony en 1914. Reposaba en una fosa oval de unos 50 cm y 40 cm de profundidad. Identificado por Bruno Maureille en 1996 en los depósitos del Museo Nacional de Prehistoria de Francia. Datado en 40,9 ka.
- L’Hortus, Languedoc. Restos de unos veinte individuos datados en 40 ka.
- Abri des Pechêurs. Cuatro dientes.
- Bouffia Bonneval, La Chapelle-aux-Saints.
- LCS 1, Old Man, descubierto por Amédée y Jean Bouyssonie en 1908. tiene una antigüedad de 40 ka y una capacidad craneal de 1.620 c.c. El esqueleto casi completo fué reconstruido por Marcellin Boule quien exageró los ragos anatómicos, popularizando el estereotípo de hombre mono. Se confundió porque no se dio cuenta que el individuo muerto a los 30 o 40 años tenía una artritis avanzada que distorsionaba sus rasgos y doblaba sus hombros. Fue enterrado intencionalmente (William Rendu et al, 2013)
- Dientes de al menos dos individuos juveniles y un adulto.
 |
| Complejo de La Chapelle aux Saints e información estratigráfica. William Rendu et al, 2013. |
 |
| Old Man. La Chapelle-aux-Saints. |
 |
| Old Man. La Chapelle-aux-Saints. |
Italia.
Grotta di Fumane. Evidencias de obtención de plumas de aves para uso simbólico.Chequia.
Dzeeraba Skala. Molar inferior. Datado con dudas en 40 ka años.Bulgaria.
Bacho Kiro. Datado en 43 ka.Croacia.
Vindija. Cráneos, mandíbula y poscraneales, resultado de prácticas caníbales. Aunque no hay duda acerca de su adscripción a neanderthalensis, la gracilidad de los restos hace sospechar de un contacto genético con el sapiens moderno. Todos los restos esqueléticos provienen del estrato G, con la posible excepción del ramus mandibular Vi-11.52 que podría pertenecer a la capa I, más antigua. Por la fauna asociada, se deduce que los neandertales habitaban la cueva en los periodos templados.- Estrato G3: Musteriense con algunos elementos del Paleolítico Superior. 43-38 ka.
- Estrato G1: Puntas de hueso Mladec y otros artefactos de hueso. Mezcla de tipos musteriense y del Paleolítico Superior. Punta de hueso Vi-3437 asociada a mandíbula Vi-207. Punta de piedra bifacial de material no-local parecida a las de la industria szeletiense.
- Interpretaciones:
- Mezcla de elementos musterienses y auriñacienses.
- Industria transicional.
- Material artificialmente mezclado (muy improbable).
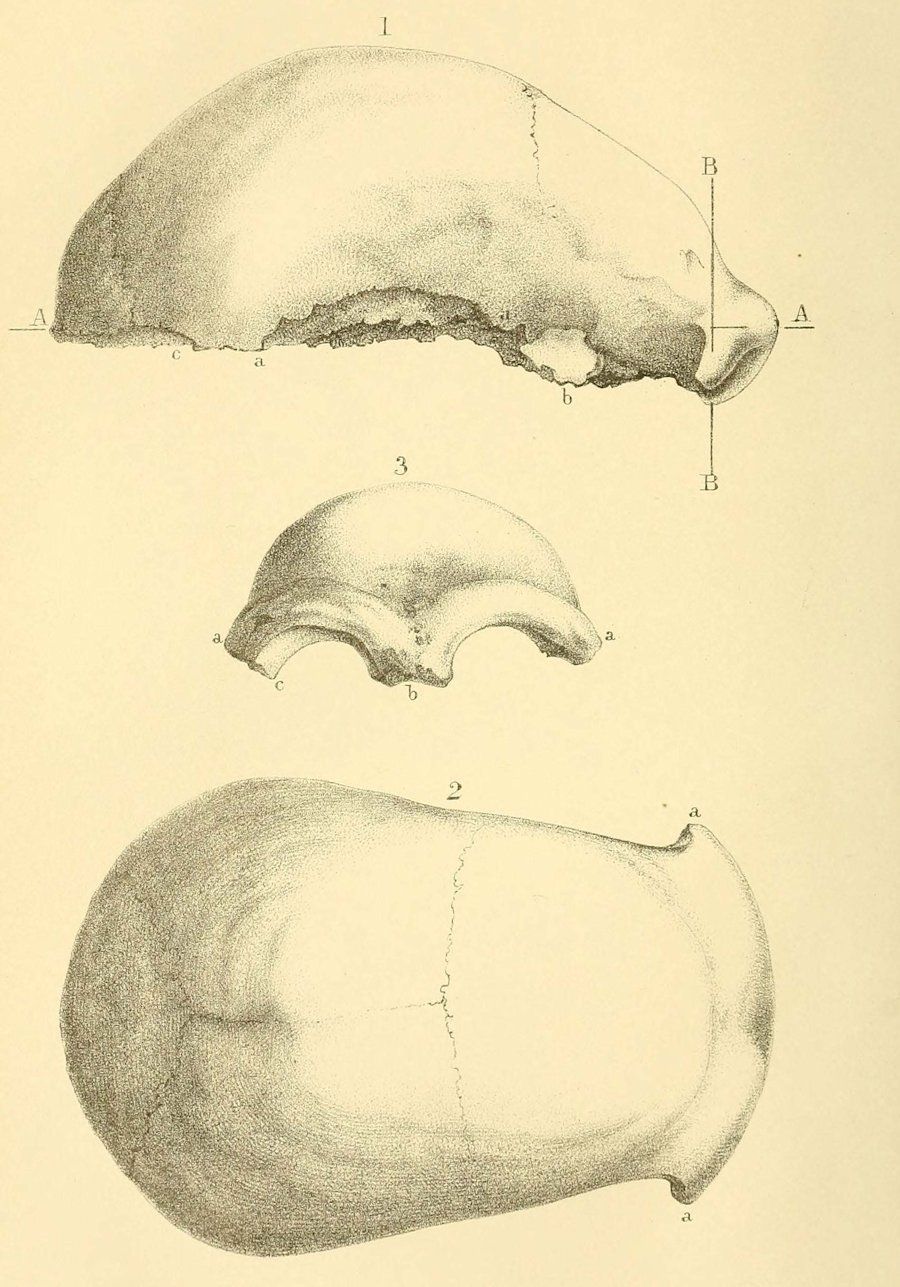 |
| Calota de Neander, parte del holotipo de Homo neanderthalensis. King, 1864. |
Alemania.
- Feldhofer, Neander. Pequeña cueva. En 1856 Johann Fuhlrott halló una bóveda craneal, huesos del fémur, parte de una pelvis, algunas costillas y huesos del brazo y hombros. El individuo al que pertenecían se rompió el antebrazo izquierdo en vida, dando como resultado que los huesos del brazo izquierdo son más cortos que el derecho.
- Königsaue, Alemania del Este. Se hallaron dos pequeños bulbos de brea de abedul, uno con una huella digital y otro con la impresión de un mango de madera. La brea de abedul es un adhesivo cuya obtención requiere un proceso técnico complejo.
Península Ibérica.
- Carihuela-1. Nivel 5.
- Arrillor. Muela infantil.
- Axlor (Indusi, Dima, Vizcaya). Maxilar incompleto y fragmento de mandíbula con varios dientes, de un mismo individuo.
- El Castillo (Puente Viesgo, Cantabria). Más de 30 fósiles de unos doce individuos.
- Banyoles. Mandíbula de adulto avanzado, con dientes muy gastados.
- Gruta da Oliveira, Almonda. 4 fragmentos.
- Cova Negra de Bellús, Xátiva, Valencia. Niveles 13-11. 2 metatarsianos, fragmento de diáfasis femoral, parietal-2.
- Calvero de la Higuera (Madrid) Lozoya, dos incisivos, un canino y una muela de una menor de 2 años y medio (2011), datados en más de 40 ka. Hay indicios de que el cuerpo de la niña fue depositado intencionalmente.
Datados en el intervalo oscilante OIS 3a (35.800-27.000)
Se constata una reducción de la robustez facial, cambios en las extremidades superiores, probablemente relacionados con el uso de la tecnología y los adornos, y en la anatomía locomotora, que denotan una forma diferente de moverse por el paisaje. Todo ello indica un cambio hacia comportamientos modernos (Trinkaus, 2011).Francia.
- Arcy-sur-Cure. Datado en 33.800 años.
- Saint-Césaire, Charente-Maritimes. Esqueleto aplastado descubierto por Francois Leveque en 1979, en un entierro secundario. El cráneo está casi completo, faltándole la parte inferior. Se ha datado alrededor de 35 ka. Se encontraron herramientas chatelperronienses, pero existe cierto grado de incertidumbre con respecto a su asociación. Poco prognatismo medio-facial, cara plana y dientes reducidos. Además, aunque no prominente, muestra una pequeña barbilla, una característica que está presente sólo en los HAM. Los arcos superciliares son muy diferentes a los de los neandertales clásicos, la frente se inclina para atrás hacia la glabela y las órbitas de los ojos y la nariz son bastante grandes.

Cráneo de Saint-Césaire

Bélgica
Gruta de Spy d' Orneau. Cráneos, mandíbulas y poscraneales (dos esqueletos casi completos Spy 1 y Spy 2) descubiertos por Marcel de Puydt y Max Lohest en 1886. Datados entre 38-34 ka. El hallazgo sirvió para desacreditar la idea de que los neandertales sufrían una especie de disminución psíquica. Los cráneos tienen entre sí diferencias significativas, lo que indica el gran margen de variación que existía dentro de la misma población. Cabe la posibilidad de que estas diferencias fueran debidas a la presencia de nuevos genes en la respectiva población.Península Ibérica
- Cueva de Gorham, Gibraltar. Forma parte de un conjunto de cuevas ocupadas por neandertales. Desde hace 125 ka, el clima fue similar al actual. Los neandertales explotaban un hábitat similar al actual de Doñana, con abundancia de presas pero también con depredadores como leones, linces, lobos y osos pardos. La costa se hallaba a unos 5 km. de la cueva. Los neandertales comían grandes mamíferos, con preferencia por la cabra montés, pero los restos más abundantes corresponden a conejos; también son muy abundantes los restos de aves. Comían también tortugas de tierra, piñones, lapas, mejillones, focas y delfines. Es de suponer que comiesen también frutas, raíces y larvas, que no dejaron rastro. Datados en 30-26 ka aparecen restos auriñacienses, lo que puede indicar la convivencia con sapiens. Las investigaciones han revelado una red de pequeñas cuevas utilizadas para las expediciones de caza ocasionalmente antes de regresar a la base.
- Devil’s Tower. Media docena de fósiles incluyendo un cráneo infantil icompleto y una mandíbula.
- Covalejos. Nivel 8.
- El Salt, Alcoi, Alicante. 6 dientes.
- Cueva de los Moros de Gabasa, Ribagorza, Huesca. Premolar, dos molares, metatarsiano, falange del pie, clavícula.
- Columbeira. Germen de primer molar.
- Figueira Brava, Sesimbra. Nivel C-2. Premolar superior.
- Salemas, Ponte de Lusa. Infantil: varios huesos y un maxilar con dientes y una muela de leche.
- Cova Negra de Bellús, Xátiva, Valencia. Niveles 9 y 8. Cráneos 3 y 5.
- Cova Foradá de Oliva. Valencia. Roca con forma de cabeza de oso cavernario hallada junto a restos humanos, con incisiones.
Últimos neandertales
En Zafarraya se ha hallado una mandíbula que muestra características neandertales clásicas, al carecer de barbilla, dientes delanteros relativamente grandes y una arcada dental grande. En 2011 Javier Baena et al comunicaron las dataciones de los
hallazgos en la Cueva del Esquilleu, Liébana, Cantabria, España. Los restos más
modernos ofrecen una datación de 20 ka. La tecnología lítica asociada
corresponde al último musteriense.
Según David Santamaría (2012) los neandertales se extinguieron
en la Península Ibérica antes de hace 40 ka.
Rachel E. Wood et al (2013) han publicado una nueva datación por el método del radiocarbono con protocolo de ultrafiltración, de los hallazgos de Cueva del Boquete de Zafarraya (Málaga) y Jarama VI (Sistema Central), dos de los últimos refugios neandertales.
En Portugal, la capa 8 de la Gruta da Oliveira proporcionó un conjunto del Paleolítico Medio (Marks et al, 2001). Se ha datado en hace 38,2-35,3 ka (Hoffmann et al, 2013).
Rachel E. Wood et al (2013) han publicado una nueva datación por el método del radiocarbono con protocolo de ultrafiltración, de los hallazgos de Cueva del Boquete de Zafarraya (Málaga) y Jarama VI (Sistema Central), dos de los últimos refugios neandertales.
La nueva datación, de 45-50ka, de diferentes ejemplares, descarta la coincidencia con Homo sapiens en estos lugares y pone en duda anteriores dataciones que asignaban 36 ka a estos reductos en la Península Ibérica. El protoauriñaciense aparece en estos lugares con dataciones de 42 ka.
También pone en duda la asignación a neandertales de herramientas y adornos en base a la datación.
Manuel Alcaraz-Castaño ha adelantado la datación del yacimiento neandertal de la Cueva de los Casares, Guadalajara, que ha deparado una antigüedad de hace 40 ka.
En Gorham’s Cave (Gibraltar) se ha documentado la presencia del Paleolítico Medio al menos hasta 37,8-36 ka (Finlayson et al, 2006; Finlayson et al, 2008). En eolianitas del Pleistoceno Tardío se han localizado huellas fósiles fechadas por OSL en 28 ± 3 ka (MIS 2). Una de estas huellas se ha asignado a Homo neanderthalensis mediante un análisis fotogramétrico (Fernando Muñiz et al, 2019).
Manuel Alcaraz-Castaño ha adelantado la datación del yacimiento neandertal de la Cueva de los Casares, Guadalajara, que ha deparado una antigüedad de hace 40 ka.
En Gorham’s Cave (Gibraltar) se ha documentado la presencia del Paleolítico Medio al menos hasta 37,8-36 ka (Finlayson et al, 2006; Finlayson et al, 2008). En eolianitas del Pleistoceno Tardío se han localizado huellas fósiles fechadas por OSL en 28 ± 3 ka (MIS 2). Una de estas huellas se ha asignado a Homo neanderthalensis mediante un análisis fotogramétrico (Fernando Muñiz et al, 2019).
En Portugal, la capa 8 de la Gruta da Oliveira proporcionó un conjunto del Paleolítico Medio (Marks et al, 2001). Se ha datado en hace 38,2-35,3 ka (Hoffmann et al, 2013).
João Zilhão et al (2017), basándose en la estratigrafía y dataciones de radiocarbono y luminiscencia han datado yacimientos de la cuenca de Mula (Murcia) concluyendo que la transición del Musteriense al Paleolítico Superior se produjo en esta región en la primera mitad del milenio 37 antes del presente:
- En Cueva Antón, la capa musteriense I-k puede ser no más antigua de hace 37.100 años.
- En el Abrigo La Boja, el Auriñaciense puede estar presente desde hace no menos de 36.500 años, en los conjuntos OH18-OH20.
Bibliografía:
- Wallace, John A., et al. "Did La Ferrassie I use his teeth as a tool? [and comments and reply]." Current'Anthropology (1975): 393-401.
- Smith, Fred H. The Neandertal remains from Krapina: a descriptive and comparative study. Diss. University of Michigan, 1976.
- Wolpoff, Milford H. "The Krapina dental remains." American Journal of Physical Anthropology 50.1 (1979): 67-113.
- Puech, Pierre-François. "Tooth wear in La Ferrassie man." Current'Anthropology 22.4(1981): 424-430.
- Wolpoff, Milford H., et al. "Upper pleistocene human remains from Vindija cave, Croatia, Yugoslavia." American Journal of Physical Anthropology 54.4 (1981): 499-545.
- Smith, Fred H., Donna C. Boyd, and Mirko Malez. "Additional upper pleistocene human remains from Vindija cave, Croatia, Yugoslavia." American Journal of Physical Anthropology 68.3 (1985): 375-383.
- Mellars, Paul. "Dating and correlating the French Mousterian (reply)." (1986): 113-114.
- Radovcic, J., et al. "The Krapina hominids: an illustrated catalog of skeletal collection." Zagreb: Croatian Natural History Museum (1988).
- Mann, Alan E., Janet M. Monge, and Michelle Lampl. "Investigation into the relationship between perikymata counts and crown formation times." American Journal of Physical Anthropology 86.2 (1991): 175-188.
- Stiner, Mary C. "The faunal remains from Grotta Guattari: a taphonomic perspective." Current'Anthropology 32.2 (1991): 103-117.
- White, Tim D., et al. "The question of ritual cannibalism at Grotta Guattari [and comments and replies]." Current'Anthropology 32.2 (1991): 118-138.
- Simek, Jan F., and Fred H. Smith. "Chronological changes in stone tool assemblages from Krapina (Croatia)." Journal of human evolution 32.6 (1997): 561-575.
- Recheisl, W., and H. Seidler. "Volume of the Guattari 1 Neandertal Specimen (Monte Circeo)." Coll.'Antropol 23.2 (1999): 397-405.
- Ahern, James, et al. "New discoveries and interpretations of hominid fossils and artifacts from Vindija Cave, Croatia." Journal of Human Evolution 46.1 (2004): 27-67.
- Thibaut Devièse, Ivor Karavanić, Daniel Comeskey, Cara Kubiak, Petra Korlević, Mateja Hajdinjak, Siniša Radović, Noemi Procopio, Michael Buckley, Svante Pääbo, and Tom Higham. 2017. Direct dating of Neanderthal remains from the site of Vindija Cave and implications for the Middle to Upper Paleolithic transition
No hay comentarios:
Publicar un comentario